Колостральний імунітет — запорука здоров’я телят

Тетяна Василенко, Андрій Кокарєв, Дмитро Масюк, НДЦ біобезпеки та екологічного контролю ресурсів АПК Дніпровського ДАЕУ
Перші години життя у новонароджених телят є одним з критичних періодів раннього постнатального онтогенезу, оскільки в цей час їх імунна система адаптується до навколишніх антигенів. Завдяки біологічним особливостям плаценти корів новонароджені телята не мають специфічних імуноглобулінів, які б забезпечували імунний захист. Саме тому передача імуноглобулінів від матері до новонародженого є життєво необхідною для телят і відбувається в перші години життя лише за допомогою молозива.
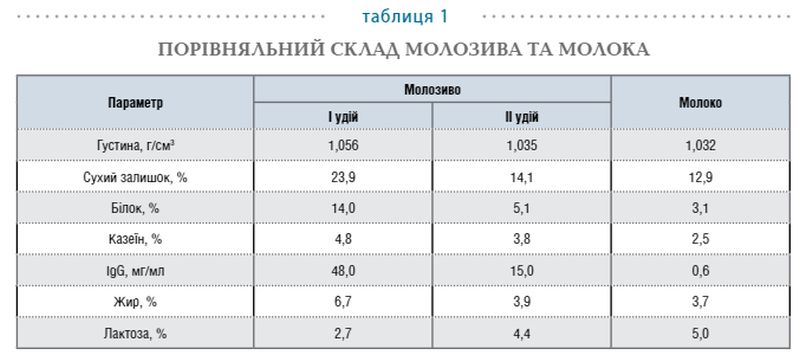
Молозиво містить усе, що потрібно молодому організмові: білки, жири, вуглеводи, мінеральні речовини, вітаміни, інші імунобіологічні речовини (табл. 1).
Завдяки унікальному складові та безпосередньо високому вмістові імуноглобулінів молозиво сприяє формуванню пасивного імунітету в новонароджених телят. Одночасно з цим молозиво володіє високим рівнем бактерицидної активності завдяки вмісту лізоциму, який розчиняє оболонки мікроорганізмів, пригнічує розвиток патогенних мікробів за рахунок високої кислотності. Тож забезпечення молозивом у перші ж години після народження надзвичайно важливе для збереження здоров'я телят і захисту їх від хвороб протягом перших тижнів життя.
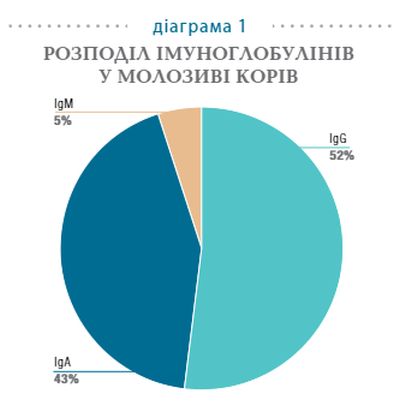 Одним із найважливіших імунобіологічних компонентів молозива є імуноглобуліни (Ig) G, А та М (діаграма 1). Вони забезпечують новонароджених телят пасивним імунним захистом. Найбільша кількість імуноглобулінів міститься в молозиві, яке секретується одразу після отелення. З часом їх кількість зменшується. Відомо, що пасивний транспорт імуноглобулінів молозива є найважливішим способом передачі імунологічного захисту. Ще у 1983 році було встановлено, що впродовж перших діб життя відбувається найактивніше поглинання імунних білків молозива у нативному (незмінному) вигляді. Після вживання молозива колостральні імуноглобуліни з’являються в периферичній крові новонароджених уже через 2 години. Це відбувається завдяки селективній абсорбції антитіл крізь кишечник, що сприяє концентрації у крові новонароджених високого рівня імуноглобулінів.
Одним із найважливіших імунобіологічних компонентів молозива є імуноглобуліни (Ig) G, А та М (діаграма 1). Вони забезпечують новонароджених телят пасивним імунним захистом. Найбільша кількість імуноглобулінів міститься в молозиві, яке секретується одразу після отелення. З часом їх кількість зменшується. Відомо, що пасивний транспорт імуноглобулінів молозива є найважливішим способом передачі імунологічного захисту. Ще у 1983 році було встановлено, що впродовж перших діб життя відбувається найактивніше поглинання імунних білків молозива у нативному (незмінному) вигляді. Після вживання молозива колостральні імуноглобуліни з’являються в периферичній крові новонароджених уже через 2 години. Це відбувається завдяки селективній абсорбції антитіл крізь кишечник, що сприяє концентрації у крові новонароджених високого рівня імуноглобулінів.
Імунологічно якісне молозиво корів містить 60−70 г/л імуноглобулінів. За даними світової статистики, нормальна кількість IgG міститься в молозиві лише у 36,4−58,6% корів, а IgМ—у 12,1−24,13%.
Імуноглобулін А, особливо його секреторна форма, відіграє важливу роль у формуванні місцевого імунного захисту. Колостральний IgA формує імунологічний бар'єр слизових оболонок кишечника в новонароджених телят з перших хвилин після споживання молозива, де у комплексі з лізоцимом секреторний IgA посилює свої бактерицидні властивості. Слід відзначити, що захисні властивості імуноглобулінів у травному каналі новонароджених проявляються лише упродовж перших 2−3 тижнів життя, що пов’язано з відсутністю вільної хлористоводневої кислоти у сичузі та відсутністю протеолізу білкових імунних молекул.
Основним імуноглобуліном молозива є IgG. До молозива він потрапляє лише з крові корів, у результаті чого різниця між його концентрацією в сироватці крові та молозиві становить близько 36%. Тому рівень колострального імунітету в крові телят, який представлений материнськими антитілами, залежить від концентрації IgG у крові, а відповідно, і в молозиві.
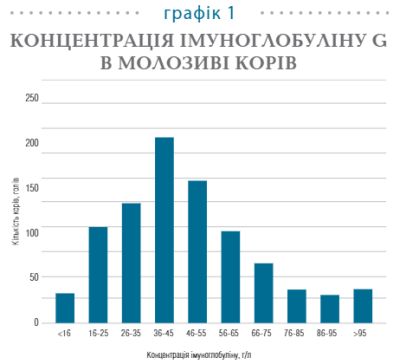 За даними низки дослідників, концентрація IgG в молозиві корів розподіляється нерівномірно, що представлено на графіку 1.
За даними низки дослідників, концентрація IgG в молозиві корів розподіляється нерівномірно, що представлено на графіку 1.
Дослідження концентрації імуноглобулінів у молозиві 919 корів виявило, що середній вміст IgG становить 48 ±22 г/л.
Основними факторами, що впливають на формування колострального імунітету, є концентрація антитіл у молозиві (що прямо корелює з біологічним станом матері), зміни природного фізіологічного стану корови (мастит перенесені захворювання під час тільності на ранньому/ пізньому терміні тощо), час випоювання молозива та його кількість, сорбційна здатність стінки кишечника теляти тощо. Максимальний рівень IgG у крові телят після народження реєструється на 5−7-у добу життя.
Слід відзначити, що тривалість колострального імунітету за рівнем загальних імуноглобулінів є різною. У середньому цей показник становить 20−30 діб (графік 2), тоді як специфічні антитіла колострального походження можуть зберігатись у крові телят 3−4 місяці.
Дослідження щодо імуноглобулінів показують, що телята 7−9-денного віку мають достатньо високий рівень загального IgG, концентрація якого в цей час коливається в межах 38−67 г/л, що в середньому становить 50,5 г/л. Це вказує на своєчасне випоювання новонародженим телятам доброякісного молозива, багатого на імуноглобуліни, що забезпечило формування в них колострального імунітету.
Дослідження концентрації IgG у сироватці крові телят віком 24−27 діб вказує на зниження його рівня в цей період, що швидше за все зумовлено природним катаболізмом колостральних антитіл. Надалі, починаючи з 35-ї доби життя, у крові телят відзначається збільшення рівня IgG, що вказує на активізацію власної імунної системи організму тварин шляхом продукції неспецифічних антитіл у відповідь на антигенні подразники зовнішнього середовища.
Отже, дослідження рівня неспецифічного IgG у сироватці крові телят протягом перших 93 діб життя дає змогу відстежити ефективність формування колострального імунітету, оцінити його тривалість та визначити період активізації власної імунної відповіді, на що вказує підвищення рівня імуноглобулінів у телят, яким більше 30 діб. Все це вельми важливо для аналізу результатів дослідження колострального імунітету за специфічними імуноглобулінами.
Як було відзначено вище, основними проблемами новонародженого молодняка ВРХ є шлунково-кишкова та респіраторна патологія, індукована впродовж перших тижнів життя тварин синергічною дією мікроорганізмів на тлі несформованого специфічного імунного захисту.
З метою недопущення виникнення інфекційної патології у телят спеціалістами ветеринарної медицини розробляються та впроваджуються в сільськогосподарських підприємствах схеми специфічної імунопрофілактики, орієнтовані на відвернення індукції інфекційних захворювань шляхом формування у тварин специфічного імунітету, що з молозивом забезпечує захист новонароджених тварин.
На сьогодні відомо, що високий рівень колостральних антитіл у крові тварин перешкоджає формуванню повноцінного імунного захисту на тлі вакцинації, оскільки колостральні антитіла мають змогу дезактивувати вакцинні антигени, тим самим істотно знижувати ефективність заходів з імунопрофілактики телят.
Тому нами було проведено дослідження колострального імунітету телят із використанням методу ELISA (ІФА) для виявлення специфічних IgG до антигенів збудників вірусної діареї, парагрипу-3 та респіраторно-синцитіальної інфекції на тлі специфічної імунопрофілактики цих хвороб у корів.
Дослідження специфічних антитіл до антигенів збудника парагрипу-3 у крові різновікових груп телят відображено на графіку 3.
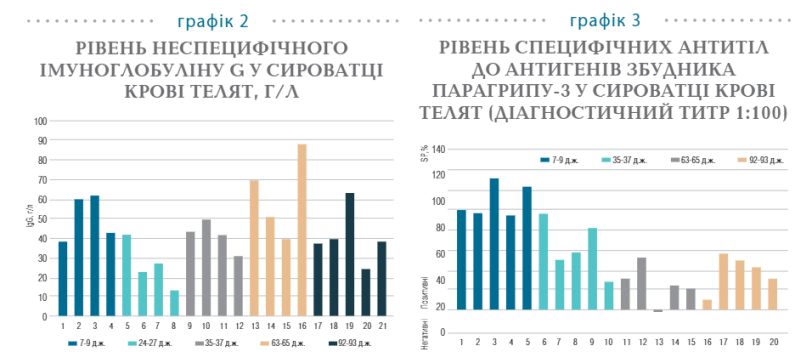
Встановлено, що всі зразки від телят 7−9-добового віку містять специфічні антитіла до антигенів вірусу парагрипу-3 із середнім значенням розрахункового коефіцієнта, який відображає рівень імуноглобулінів (SP)—101%. Надалі, на 35−37-у добу життя, за рівнем SP спостерігається зменшення вмісту цих антитіл у крові телят до 68%, а на 63−65-у добу набуває найменшого значення—39%. Слід відзначити, що на тлі низхідної динаміки рівня антитіл усі тварини 35−37-добового віку є серопозитивними, що свідчить про повноцінно сформований груповий імунітет, тоді як на 63−65-у добу один із п’яти зразків є серонегативним за парагрипом-3 у діагностичному титрі 1:100. Присутність у стаді неімунних телят, або тварин з низьким титром антитіл, сприяє активації вищезазначеної інфекції у стаді та, як наслідок, збільшення пресингу вірусу.
На 92−93-ю добу життя помічено збільшення рівня специфічних антитіл у крові телят, що позначається підвищенням середнього значення по групі SP до 49%. Така динаміка імуноглобулінів пояснюється сероконверсією антитіл в організмі тварин, що позитивно реагують на дію антигенів до збудника парагрипу-3. Схожі результати було отримано під час дослідження специфічних антитіл до антигенів збудника респіраторно-синцитіальної інфекції (графік 4).
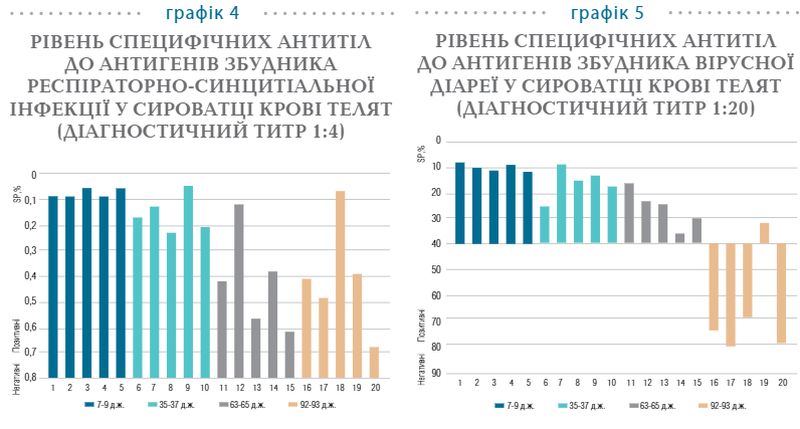
Всі досліджувані групи телят мають на 100% сформований груповий імунітет до респіраторно-синцитіальної інфекції, який у перші 63−65 діб представлений специфічними антитілами колострального походження, на що вказує їх поступове, але відчутне зниження впродовж цього періоду. Надалі відзначається збільшення рівня імуноглобулінів, що, як і у випадку із парагрипом-3, зумовлено сероконверсією антитіл організмом тварин, що позитивно реагують на дію антигенів респіраторно-синцитіального вірусу. Остання може відбуватись унаслідок інфікування тварин епізоотичним штамом вірусу, що циркулює у стаді, або на тлі вакцинації тварин, яка відбулась у 2-місячному віці. Результати дослідження колостральних антитіл, специфічних до антигенів збудника вірусної діареї ВРХ, вказують на стопроцентно сформований груповий імунітет у телят до 63−65-добового віку (графік 5). Рівень специфічних імуноглобулінів поступово знижується впродовж 63−65 діб життя, про що свідчить підвищення коефіцієнта SP, який має обернено пропорційну залежність. На 92−93-ю добу життя 80% досліджуваних проб є серонегативними за вірусною діареєю, що позначилось високим значенням коефіцієнта SP у діагностичному титрі 1:20. Це вказує на формування «серологічного вікна», яке може сприяти інфікуванню серонегативних телят збудником вірусної діареї ВРХ.
Таким чином, колостральний імунітет до вірусної діареї, респіраторно- синцитіальної інфекції та парагрипу-3 у телят є повноцінно сформованим та зберігається на високому рівні упродовж перших 30−35 діб життя, після чого його рівень поступово зменшується, що пов’язано з фізіологічним катаболізмом материнських антитіл, і вже з 65-ї доби від народження виявляються тварини, які є серонегативними.
Підсумовуючи вищенаведене, можна зробити висновок, що оцінювання ефективності формування неспецифічного колострального імунітету необхідно проводити на 7−9-у добу життя за показником загального (неспецифічного) IgG, концентрація якого в цей час повинна становити не менше 10 г/л.
Зважаючи на те, що тривалість колострального специфічного імунітету в телят не відповідає загальній динаміці рівня неспецифічних материнських антитіл і з метою виявлення періоду формування «серологічного вікна», який сприяє інфікуванню тварин наявними у стаді вірусами, для формування або коригування схеми лікувально-профілактичних заходів у телят необхідно проводити дослідження рівня специфічних антитіл до вірусних антигенів протягом перших 90 діб життя, починаючи з 7-денного віку, з інтервалом між віковими групами для дослідження не більше, ніж 14−21 доба.
Джерело: журнал «Молоко і ферма» № 2 (63) квітень, 2021